车明晓 张新节 陆梓晔 迟淑艳,3,4* 谭北平,3,4
(1.广东海洋大学水产学院,水产动物营养与饲料实验室,湛江524088;
2.湛江国联饲料有限公司,湛江524051;
3.广东省水产动物精准营养与高效饲料工程技术研究中心,湛江524088;
4.农业农村部华南水产与畜禽饲料重点实验室,湛江524088)
糖类是动物最经济的能量来源,鱼类饲料中添加适量的糖类可以起到节约蛋白质的作用[1-2]。然而,鱼类对糖类的利用能力因种类而异[3],并且受饲料中糖类水平和结构的影响[4-9]。糖类按照结构可分为单糖、双糖、低聚糖和多聚糖等。蔗糖是由葡萄糖和果糖组成的双糖,而多糖中的淀粉则是目前鱼类饲料中使用最为广泛的糖类来源[10]。研究表明,肉食性鱼类对多糖的利用能力要强于单糖和双糖[6-7,11];
与多糖相比,单糖和双糖更容易被肠道消化和吸收[3,12],使得餐后血糖迅速上升,但肉食性鱼类清除血糖速度较杂食性鱼类和草食性鱼类慢[13]。通常,鱼对饲料的表观消化率和肠道吸收率随着摄食的糖类物质结构复杂性的增加而降低[9],而白鲟(Psephurusgladius)[8]利用葡萄糖和和麦芽糖的能力比果糖、蔗糖、乳糖、糊精或淀粉都强,因此选择适当的糖类来源,以最大限度地减少蛋白质分解代谢的能量至关重要。
大口黑鲈(Micropterussalmoides)是一种典型的肉食性经济鱼类,利用糖类的能力差[14]。本试验探究了不同糖源对大口黑鲈生长性能、血清生化指标和肝脏糖脂代谢等方面的影响,旨在为该物种提供更有效的饲料原料。
1.1 试验饲料
以红鱼粉、鸡肉粉和酪蛋白为主要蛋白质源,大豆油和鱼油为主要脂肪源,分别以葡萄糖(Glu组)、蔗糖(Suc组)、糊精(Dex组)、玉米淀粉(CS组)、豌豆淀粉(PS组)和木薯淀粉(TS组)为糖源,配制6种等氮等脂的试验饲料(其组成及营养水平见表1)。所有试验饲料均由广东粤海饲料集团股份有限公司提供。将原料粉碎过60目筛网,按照配方准确称取,采用逐级扩大法将原料混合均匀,在V型混合机(B30,广州市番禺力丰食品机械厂)中混合12 min,然后加入鱼油、大豆油和水(25%,质量分数),用搅拌机搅拌均匀后,使用干法膨化机制作沉性膨化颗粒饲料,模板孔径为3 mm,饲料颗粒长度为3 mm,制粒完成后在25 ℃空调房中自然晾干,晾至水分含量约为10%后装入封口袋,存放于-20 ℃的冰箱中。

表1 试验饲料组成及营养水平(饲喂基础)

续表1项目 Items组别 GroupsGluSucDexCSPSTS糊精 Dextrin11.0玉米淀粉 Corn starch11.0豌豆淀粉 Pea starch11.0木薯淀粉 Tapioca starch11.0微晶纤维素 Microcrystalline cellulose4.94.94.94.94.94.9多维预混料 Multi-vitamin premix2)1.01.01.01.01.01.0多矿预混料 Multi-mineral premix2)3.03.03.03.03.03.0合计 Total100.0100.0100.0100.0100.0100.0营养水平 Nutrient levels水分 Moisture10.119.8710.3410.029.719.96粗蛋白质 CP50.0450.2549.8349.7750.3550.16粗脂肪 EE10.1510.3110.1910.2610.4610.33可消化碳水化合物 Digestible carbohydrates11.0611.2110.9511.1410.9911.11粗灰分 Ash12.5512.9012.6813.1112.7313.26
1.2 试验设计和饲养管理
试验选择遗传背景一致的大口黑鲈鱼苗(高州市泗水镇英熙鱼虾苗场),在桶中暂养2周,暂养期间投喂商品饲料(粗蛋白质48%、粗脂肪7%)。正式试验开始前,禁饲24 h,挑选健康、规格均匀的鱼苗540尾[初始体重(5.01±0.04) g],随机分配到18个0.4 m3的玻璃纤维钢化桶中,分为6组,每组3个重复,每桶30尾鱼苗。每天08:00和16:00投喂至表观饱食。养殖试验在广东省湛江国联饲料有限公司室内循环水养殖系统进行,溶解氧含量>5 mg/L,水温28~31 ℃。试验期56 d。
1.3 样品采集及分析
1.3.1 生长性能指标
在试验结束时,禁食24 h,对每个桶的鱼进行计数和称重,以计算存活率(SR)、增重率(WGR)和特定生长率(SGR);
在喂养过程中记录饲料摄入量,并用于计算摄食率(FI)和饲料系数(FCR)。
从每个桶中随机取5条鱼测量体长和体重,然后解剖鱼体,剥离并称量内脏团和肝脏,用以计算肥满度(CF)、肝体比(HSI)和脏体比(VSI)。
各生长性能指标用以下公式计算:
存活率(%)=100×试验结束时每桶鱼的数量/
试验开始时每桶鱼的数量;
增重率(%)=100×(鱼末重-鱼初重)/鱼初重;
特定生长率(%/d)=100×(ln鱼末重-
ln鱼初重)/试验天数;
饲料系数=每尾鱼的摄食量/(鱼末重-鱼初重);
摄食率(%/d)=100×每尾鱼的摄食量/
[试验天数×(鱼末重+鱼初重)/2];
肥满度(g/cm3)=100×体重/体长3;
肝体比(%)=100×肝脏重量/体重;
脏体比(%)=100×内脏团重量/体重。
1.3.2 饲料营养成分测定
饲料营养成分采用AOAC(1997)方法进行分析。水分测定采用105 ℃恒重干燥法,粗蛋白质测定采用凯氏定氮法(KjeltecTM8400,瑞典,N×6.25),粗脂肪测定采用索氏提取法(抽提剂为石油醚,沸点为30~60 ℃)。
1.3.3 血清生化指标
3.2 国内外同类研究比较 国内外对先天性心脏病患儿神经心理发育水平的研究多以重症或复杂先天性心脏病为主[3],其表现有不同程度的心理发育迟滞和智力落后,语言发育迟缓,甚至出现脑瘫、癫痫等并发症[4],其原因多归因于心脏病理改变所致的胎儿期、出生后、手术中及术后各期的脑和神经系统的缺血缺氧性损伤[5]。国内先心病儿童生长发育的研究则多以身长体重等体格生长发育指标的评估及影响因素分析为主。国内外对本研究涉及的病情较轻的小型室间隔缺损患儿的神经心理发育研究相对少见,可比性资料不多,一般观点均以患儿神经心理发育水平大致处于正常范围加以概括性的推测,缺少实证性研究。
从每个桶中随机抽取5条鱼,用1 mL的无菌注射器进行尾部静脉抽血,将血样转移到1.5 mL离心管中,在4 ℃下保存12 h,在4 ℃下以3 500 r/min离心15 min,获得血清。血清中的甘油三酯(TG,货号A110-1-1)、总胆固醇(TC,货号A111-1-1)、高密度脂蛋白胆固醇(HDL-C,货号A112-1-1)、低密度脂蛋白胆固醇(LDL-C,货号A113-1-1)和葡萄糖(货号A154-1-1)含量及谷草转氨酶(AST,货号C010-2-1)和谷丙转氨酶(ALT,货号C009-2-1)活性使用试剂盒(南京建成生物工程研究所)经酶标仪(Multiskan,Thermo,美国)测量;
血清胰岛素(INS,货号ml290818)含量采用酶联免疫吸附试验(ELISA)试剂盒(上海酶联生物科技有限公司)测定。
1.3.4 肝脏酶活性及糖原和肌糖原含量测定
随机取出3条鱼的肝脏和肌肉分别置于冻存管中,用液氮保存,然后转移至-80 ℃保存。称取2 g的肝脏或肌肉组织,使用匀浆介质以1∶9比例于4 ℃下对组织进行研磨、稀释,然后于4 ℃下4 000 r/min离心15 min,取上清液。肝脏中葡萄糖激酶(GK,货号ml024808)、磷酸果糖激酶(PFK,货号ml258084)、丙酮酸激酶(PK,货号ml954600)、磷酸烯醇丙酮酸羧化激酶(PEPCK,货号ml036430)、葡萄糖-6-磷酸酶(G-6-Pase,货号ml036419)、脂肪酸合成酶(FAS,货号 ml036370)、激素敏感性脂肪酶(HSL,货号ml026143)和肉碱棕榈酰转移酶1(CPT1,货号ml036411)活性采用试剂盒(上海酶联生物科技有限公司)经酶标仪(Multiskan,Thermo,美国)测定。肝脏和肌肉糖原(货号A043-1-1)含量、肝脏丙二醛(MDA,货号A003-1-2)含量以及过氧化氢酶(CAT,货号A007-1-1)、谷胱甘肽过氧化物酶(GSH-PX,货号A005-1-2)和超氧化物歧化酶(SOD,货号A001-3-2)活性采用试剂盒(南京建成生物工程研究所)经酶标仪(Multiskan,Thermo,美国)测定。
1.3.5 肝脏切片
随机取出2条鱼的肝脏置于4%甲醛溶液中保存,委托武汉塞维尔生物技术有限公司制作切片。
1.3.6 RNA提取、反转录及mRNA定量分析
每个桶随机取3条鱼,采集肝脏样品,用生理盐水清洗后,放入含有适量RNAlater(Ambion,美国)的无酶EP管中,在-80 ℃下保存。用TRIzol试剂(北京全式金生物技术股份有限公司)从大口黑鲈肝脏中提取总RNA,用1.0%变性琼脂糖凝胶电泳检测其完整性。用NanoDrop 2000(Thermo Fisher Scientific,美国)测定RNA的纯度和浓度。合格的RNA用无RNA的DNase试剂处理以去除DNA污染物,并使用分光光度计(ND-1000,Nano-Drop Technologies,Wilmington,美国)评估RNA的提取质量。用PrimeScript®RT-PCR试剂盒(TaKaRa,日本)反转录成cDNA。
通过实时荧光定量PCR测定大口黑鲈肝脏中相关基因的mRNA表达。根据本实验室的大口黑鲈转录组数据,设计肝脏代谢酶序列的特定引物(表2)。以延伸因子1α(EF1α,GenBank登录号,XM_038724777.1)作为内参基因,根据SYBR®Premix Ex TaqTMⅡ试剂盒说明书进行PCR反应。实时荧光定量PCR反应体系为20 μL,包括10 μL 2×ChamQ SYBR qPCR Master Mix、0.4 μL PCR正向引物(10 μmol/L)、0.4 μL PCR 反向引物(10 μmol/L)、4 μL cDNA模板和5.2 μL双蒸水。PCR反应条件为95 ℃预变性70 s,95 ℃变性5 s,60 ℃退火15 s,72 ℃延伸20 s,40个循环。每个样品重复进行3次,使用2-△△Ct方法计算目标基因的mRNA相对表达量。

表2 实时荧光定量PCR引物序列

续表2基因Genes引物序列 Primer sequences (5"—3")产物长度Produce length/bp退火温度 Annealing temperature/℃GenBank登录号GenBank accession number磷酸肌醇-3-激酶调节亚基1 PI3KR1F:GTGAGGACCCGGATAAATACACR:TTCACCTGCCCACAGTAAAG17661.6XM_038718511.1蛋白激酶B AktF:ATGGACTCCTCTCCAGACCCR:TTCATGGCGTAGTAGCGTCC21360.5XM_038729214.1磷酸烯醇丙酮酸羧化激酶 PEPCKF:TCCATCCATCGTCAACCGCTTAR:ACACCGCCATCGCTAGTCTCT20558.5XM_038725393.1葡萄糖-6-磷酸酶催化亚基 G6PCF:GCAACAAATCCACCAACAAGGR:CAGGAGACCCAAAGAAACAAGC16360.6XM_038735542.1延伸因子1α EF1αF:TGCTGCTGGTGTTGGTGAGTTR:TTCTGGCTGTAAGGGGGCTC23356.8XM_038724777.1
1.4 数据统计分析
试验数据采用SPSS 21.0进行单因素方差分析(one-way ANOVA),并用Duncan氏法进行多重比较,结果以“平均值±标准差(mean±SD)”表示,P<0.05表示差异显著。
2.1 饲料糖源对大口黑鲈生长性能的影响
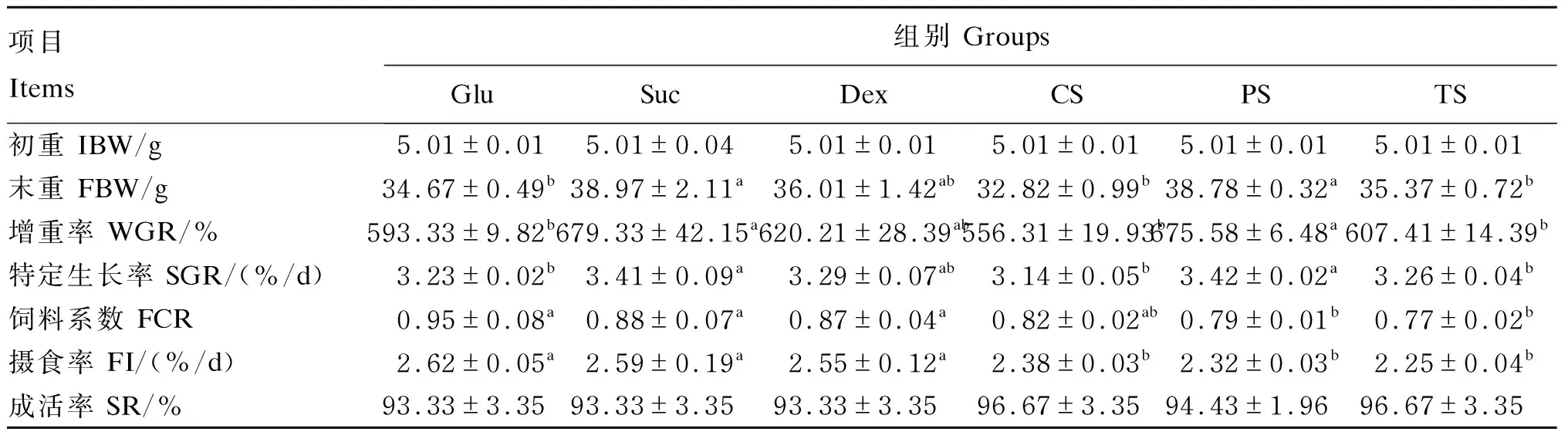
表3 饲料糖源对大口黑鲈生长性能的影响
2.2 饲料糖源对大口黑鲈形态学指标的影响
饲料糖源对大口黑鲈形态学指标的影响见表4。由表可知,TS组的肥满度显著高于Dex组(P<0.05),与其他组无显著差异(P>0.05);
Dex组和CS组的肝体比显著高于Glu组和Suc组(P<0.05),与PS组和TS组无显著差异(P>0.05);
Suc组的脏体比显著低于其他组(P<0.05)。

表4 饲料糖源对大口黑鲈形态学指标的影响
2.3 饲料糖源对大口黑鲈血清生化指标的影响
饲料糖源对大口黑鲈血清生化指标的影响见表5。由表可知,Suc组血清TG含量显著高于其他组(P<0.05),且Dex组、CS组、PS组和TS组之间无显著差异(P>0.05),但均显著低于Glu组(P<0.05);
CS组和TS组血清TC含量显著低于其他组(P<0.05);
PS组血清HDL-C含量显著高于Suc组、Dex组和TS组(P<0.05);
PS组和TS组血清LDL-C含量显著低于其他组(P<0.05);
Dex组、CS组、PS组和TS组血清AST和ALT活性显著高于Glu组和Suc组(P<0.05),且CS组血清ALT活性显著高于其他组(P<0.05);
Glu组血清葡萄糖含量显著高于Suc组和PS组(P<0.05),与其他组无显著差异(P>0.05);
Glu组和CS组血清胰岛素含量显著高于其他组(P<0.05),且Suc组和PS组显著低于其他组(P<0.05)。

表5 饲料糖源对大口黑鲈血清生化指标的影响
2.4 饲料糖源对大口黑鲈肝脏和肌肉糖原含量的影响
饲料糖源对大口黑鲈肝脏和肌肉糖原含量的影响见表6。由表可知,Suc组、Dex组和TS组肝糖原含量显著高于Glu组和PS组(P<0.05);
Glu组、PS组和TS组肌糖原含量显著高于Suc组、Dex组和CS组(P<0.05)。

表6 饲料糖源对大口黑鲈肝脏和肌肉糖原含量的影响
2.5 饲料糖源对大口黑鲈肝脏组织形态的影响
饲料糖源对大口黑鲈肝脏组织形态的影响见图1。由图可知,Glu组和Suc组肝脏细胞排列不规则,大小不均,细胞较其他组小,细胞界限模糊,细胞膜破裂,一些细胞核聚集在一起;
而Dex组、CS组、PS组和TS组肝脏细胞大小较为均一,细胞核向细胞膜靠近,有轻微细胞空泡化现象。

红色箭头:细胞膜破裂,细胞轮廓模糊;
黄色箭头:细胞空泡化;
黑色箭头:细胞核偏移。
2.6 饲料糖源对大口黑鲈肝脏抗氧化指标的影响
饲料糖源对大口黑鲈肝脏抗氧化指标的影响见表7。由表可知,PS组肝脏CAT、GSH-Px和SOD活性显著高于Glu组(P<0.05),肝脏MDA含量显著低于Glu组、Suc组和TS组(P>0.05);
Glu组肝脏CAT活性显著低于Suc组、CS组、PS组和TS组(P<0.05),与Dex组无显著差异(P>0.05);
Suc组肝脏GSH-Px活性显著低于其他组(P<0.05),Glu组、Dex组和TS组之间无显著差异(P>0.05),CS组显著高于Suc组(P<0.05),并显著低于其他组(P<0.05);
Glu组和Suc组肝脏MDA含量显著高于其他组(P<0.05);
CS组、PS组和TS组肝脏SOD活性较高,显著高于Glu组(P<0.05)。

表7 饲料糖源对大口黑鲈肝脏抗氧化指标的影响
2.7 饲料糖源对大口黑鲈肝脏糖代谢酶活性的影响
饲料糖源对大口黑鲈肝脏糖代谢酶活性的影响见表8。由表可知,Dex组肝脏GK活性显著高于Glu组、CS组和PS组(P<0.05),且PS组显著高于Glu组和CS组(P<0.05),与Suc组和TS组无显著差异(P>0.05);
Suc组肝脏PFK活性显著低于CS组和TS组(P<0.05);
Dex组、PS组和TS组肝脏PK活性显著高于其他组(P<0.05),且Suc组显著低于其他组(P<0.05);
PS组肝脏G-6-Pase活性显著高于其他组(P<0.05),且Suc组显著低于PS组和TS组(P<0.05),Glu组和Dex组显著低于其他组(P<0.05);
PS组肝脏PEPCK活性显著高于Glu组、Suc组、Dex组和TS组(P<0.05),与CS组无显著差异(P>0.05),且Dex组显著高于Suc组(P<0.05),并与Glu组无显著差异(P>0.05)。

表8 饲料糖源对大口黑鲈肝脏糖代谢酶活性的影响
2.8 饲料糖源对大口黑鲈肝脏脂代谢酶活性的影响
饲料糖源对大口黑鲈肝脏脂代谢酶活性的影响见表9。由表可知,Suc组肝脏FAS活性显著高于Glu组(P<0.05),与其他组无显著差异(P>0.05);
PS组肝脏CPT1活性显著高于Glu组和Suc组(P<0.05),肝脏HSL活性显著高于Glu组、Suc组和Dex组(P<0.05),与其他组无显著差异(P>0.05)。

表9 饲料糖源对大口黑鲈肝脏脂代谢酶活性的影响
2.9 饲料糖源对大口黑鲈糖代谢及胰岛素途径相关基因mRNA相对表达量的影响
饲料糖源对大口黑鲈糖代谢及胰岛素途径相关基因mRNA相对表达量的影响见图2。由图可知,Dex组、CS组、PS组和TS组肝脏GKmRNA相对表达量显著高于Glu组和Suc组(P<0.05),且Suc组显著高于Glu组(P<0.05);
CS组、PS组和TS组肝脏胰岛素受体(INSR)mRNA相对表达量显著高于Glu组和Suc组(P<0.05),且PS组显著高于Dex组和CS组(P<0.05);
Glu组肝脏胰岛素受体底物(IRS)mRNA相对表达量显著低于其他组(P<0.05),且PS组显著高于Suc组和TS组(P<0.05);
Dex组、CS组、PS组和TS组肝脏磷酸肌醇-3-激酶调节亚基1(PI3KR1)、蛋白激酶B(Akt)和PEPCKmRNA相对表达量显著高于Glu组和Suc组(P<0.05),且PS组显著高于其他组(P<0.05);
CS组、PS组和TS组肝脏葡萄糖-6-磷酸酶催化亚基(G6PC)mRNA相对表达量显著高于Glu组、Suc组和Dex组(P<0.05),且PS组显著高于CS组(P<0.05)。

数据柱标注相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05)。
本试验结果表明,饲料不同糖源显著影响大口黑鲈的生长性能和饲料利用率。摄食葡萄糖的大口黑鲈增重率和特定生长率均较低,并伴随着饲料系数的升高,说明大口黑鲈对葡萄糖的利用能力差。单糖对大口黑鲈生长性能的影响与团头鲂(Megalobramaamblycephala)[15]、吉富罗非鱼(Oreochromisniloticus)[16]、异育银鲫(Carassiusauratusgibelio)[17]、金头鲷(Sparusaurata)[18]、军曹鱼(Rachycentroncanadum)[19]和长吻鮠(Leiocassislongirostris)[17]中的研究结果一致。据报道,当大西洋鲑鱼(Salmosala)、红鲷(Beryciformes)、罗非鱼和白鲟的血糖水平升高时,过多的葡萄糖可能会从尿液或鳃中排出,在喂食高糖饲料中观察到有葡萄糖尿的现象[20],而在饲料中添加淀粉时,葡萄糖排泄量很低或为零,并且随着糖类物质分子复杂性的降低而增加[21]。同时,严重的美拉德反应会降低饲料中营养物质的利用率[21],这可能是饲喂葡萄糖导致生长性能下降的原因之一。
在喂食高糖饲料后,胰岛素作为一种合成代谢激素加强了葡萄糖的吸收[22]。摄食单糖饲料的大口黑鲈血清胰岛素含量显著高于摄食淀粉组,并伴有明显的高血糖,这表明摄食单糖会诱导大口黑鲈胰岛素抵抗,因此,其生长性能、成活率和饲料利用率较低,可能与糖代谢紊乱有关。单糖比多糖更容易吸收,更容易引起高血糖[23],持续性高血糖可显著促进代谢紊乱并导致代谢综合征[24-25]。相较于淀粉组,葡萄糖组的糖酵解酶活性降低,葡萄糖分解代谢能力减弱,虽然糖异生酶活性同样降低,但葡萄糖无法分解利用,依然促使高血糖的持续发生[26],这加剧了葡萄糖代谢紊乱,导致大口黑鲈生长减慢。
对大黄鱼(Larimichthyscrocea)[17]、克林雷氏鲶(Rhamdiaquelen)[11]和斑点叉尾(Ictaluruspunctatus)[7]进行的研究表明,与饲料淀粉相比,饲料中的蔗糖也显著降低了增重率、特定生长率、饲料效率和蛋白质效率。而本试验中,摄食蔗糖饲料和摄食豌豆淀粉饲料的大口黑鲈在增重率、特定生长率、血清葡萄糖和胰岛素含量方面没有显著差异,但前者饲料系数显著提高,这表明大口黑鲈可以在短期内通过稳态调节机制来适应饲料中的蔗糖,但无法很好地利用。
通常,营养代谢与肝损伤之间的关系密切,肝脏损伤通常伴随着肝脏重量和肝体比的增加[27]。对澳洲宝石鲈(Scortumbarcoo)[28]、牙鲆(Paralichthysolivaceus)[29]和异育银鲫[17]的研究也报告了肝脏中糖原的增加伴随着较高的肝体比。摄食葡萄糖和蔗糖导致肝脏损伤,但肝脏损伤相关指标的水平反而降低,这种“损害救济”可能不是一种正常状态。本试验研究结果显示,摄食葡萄糖和蔗糖的大口黑鲈血清AST和ALT活性显著低于饲喂淀粉饲料的大口黑鲈。此外,机体糖原含量也受到饲料糖源的影响[30],本试验中大口黑鲈摄食葡萄糖和蔗糖,肝脏中的糖原含量也较低,这可能也是导致肝体比下降的重要原因之一,而PS组肝糖原含量明显低于CS组和TS组,表明摄食豌豆淀粉可以减少肝脏糖原蓄积。与罗非鱼[31]可以将蔗糖通过增强肌肉糖异生能力转换成肌糖原不同的是,大口黑鲈摄食蔗糖后肌糖原含量低于淀粉组,而血清TG含量和肝脏FAS活性含量明显升高,这表明与罗非鱼相比,大口黑鲈摄食蔗糖后更多的将其转换为脂肪。
本试验中,与Glu组相比,PS组大口黑鲈的增重率和特定生长率显著提高,饲料系数显著降低,而CS组和TS组则与Glu组在生长性能方面没有显著差异,但饲料系数显著降低。使用淀粉为糖源可以显著降低血清葡萄糖和胰岛素含量,其原因可能是由于消化酶活性以及多糖要分解为葡萄糖进入血液,存在吸收的时间差[20]。淀粉的消化速度慢,有助于减少摄食淀粉后造成的葡萄糖应激[20,32],降低胰岛素峰值分泌[33];
而且,淀粉还存在直链淀粉和支链淀粉,本试验中,摄食率没有因淀粉来源不同而发生显著改变,而PS组增重率最高,这可能是因为豌豆淀粉中直链淀粉含量最高,而抗性淀粉的生成与淀粉中直链淀粉的比例呈正相关[34],因此,高直支比的淀粉导致淀粉的缓慢消化,这可能有助于改善鱼类的生长性能[35-36]。
在暗纹东方鲀(Takifuguobscurus)的研究中观察到,直链淀粉与支链淀粉的比例提高导致其GK活性显著降低[36],欧洲鲈鱼(Dicentrarchuslabrax)则相反[37]。GK催化了葡萄糖利用的第一步[38],摄食糖类物质会明显促进GK的活性和基因表达。在本试验中,不同淀粉来源对大口黑鲈肝脏GKmRNA相对表达量无显著影响,但PS组的肝脏GK活性要显著高于CS组,表明摄食豌豆淀粉可以促进大口黑鲈对葡萄糖的利用。
在哺乳动物中,胰岛素途径在调节葡萄糖代谢以避免肝糖原积累方面起着重要作用[39]。同时,饲料中的糖源可以显著影响胰岛素途径[40]。在本试验中,大口黑鲈肝脏INSR、IRS和PI3KR1的mRNA相对表达量被豌豆淀粉明显上调,这表明与其他处理相比,豌豆淀粉在转录水平上胰岛素途径的活性很高。一般来说,胰岛素途径的激活通常会导致哺乳动物体内糖酵解的上调和糖异生的下调[39]。然而,在本研究中,豌豆淀粉改善了糖酵解,而糖异生却没有受到抑制。
本试验条件下:
① 大口黑鲈摄食蔗糖可达到与摄食豌豆淀粉相当的生长性能,但饲料利用效率和抗氧化能力较低;
② 与玉米淀粉和木薯淀粉相比,摄食豌豆淀粉更有利于大口黑鲈的生长、饲料效率、抗氧化能力和对糖的利用。
猜你喜欢大口货号摄食鸡内金打粉 让孩子大口大口地吃饭基层中医药(2021年6期)2021-11-02鞋品牌新品爆单“故事汇”服饰导报·鞋世界(2021年4期)2021-05-17两种不同投喂策略对加州鲈摄食量和生长的影响当代水产(2021年1期)2021-03-19轻则摄食减慢,重则大量死鱼!加州鲈养殖亚硝酸盐超标,预防处理如何做好?当代水产(2019年9期)2019-10-08笼养灰胸竹鸡入冬前日摄食量规律的研究甘肃畜牧兽医(2019年2期)2019-03-20作者更正致歉说明中国药理学通报(2019年5期)2019-01-11大口小口话水果创新作文(小学版)(2018年10期)2018-08-20大口黑鲈饲料营养需求研究现状广东饲料(2016年7期)2016-12-01摄食紫菜粉等4种单一饵料的蚤状溞脂肪酸组成的比较应用海洋学学报(2014年1期)2014-11-22大学生网球装备推荐网球俱乐部(2009年5期)2009-06-15

